
by Connor B. Driscoll, Tim G. Otten, Theo W. Dreher
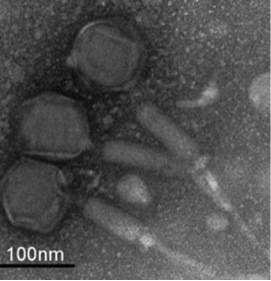
Figure 1. Microcystis virus Ma-LMM01 (Yoshida, et al. 2007)
Microcystis is a harmful algal bloom that forms cyanobacteria that can produce a potent hepatotoxin known as microcystin. The full range of factors that could affect Microcystis bloom collapse is poorly understood (with the role of virus notably lacking). In addition, our lab has shown that Microcystis strain successions often occur within the span of a single bloom, but the causes for strain decline or turnover are not understood (Bozarth, 2010). The identification of a Microcystis virus known as Ma-LMM01, which infects Microcystis aeruginosa strain NIES-298, highlights the possibility that viruses may be responsible for some cases of bloom collapse or strain turnover (Fig. 1, Yoshida, et al. 2007).
To explore this, for twelve weeks in 2011 and 2012 surface grab samples were collected from a Microcystis bloom at various sites in the San Francisco Delta at two-week intervals. Samples were put through a 0.2 µm filter, allowing free virus to pass through but keeping bacteria on the filter. Total DNA from these filters was extracted, and quantitative polymerase chain reaction (QPCR) for Microcystis marker genes was used to quantify the Microcystis population from each site.
Figure 2. Mildred Island in the San Francisco Delta, near Stockton, CA.
We chose to focus on the data collected from Mildred Island (Fig. 2), because of unique changes observed in the Microcystis population there. After a steady increase in Microcystis abundance, there was a rapid decline between samples collected on 8/27/12 and 9/10/12 (Fig. 3). This contrasts with standard bacterial growth curves, which include a stationary phase between the growth and death phases (Fig. 4). While we may have missed this phase due to sampling time, there were no dramatic population changes at other Delta sites, and no significant changes in the environmental factors measured. This could suggest that the 2012 Mildred Island Microcystis bloom was a virus-mediated bloom collapse.
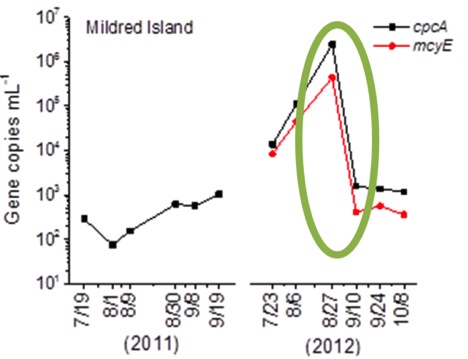
Figure 3. QPCR of Mildred Island sample. Green circle highlights bloom decline between 8/27 and 9/10.
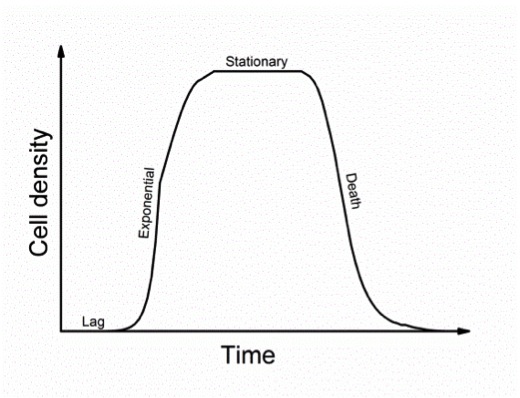
Figure 4. Bacterial Growth Curve.
Figure 5. Sequencing assembly.
To further explore this possibility, DNA extracted during bloom (8/27/12) and after bloom collapse (9/10/12) was submitted for metagenome sequencing. This method produces millions of short DNA sequence reads which can be searched for their similarity to organisms of interest, as well as assembled together into longer sequences via overlapping reads (Fig. 5). As expected, initial searches revealed a high number of reads similar to Microcystis NIES-843 in the bloom dataset and a massive drop in number of similar reads from the collapse dataset (Table 1). Interestingly, parts of 12 putative phage genomes were identified in assembled sequences. Their presence indicated that these viruses were actively infecting cells collected on the filters. One of these viruses strongly resembled the above-mentioned virus Ma-LMM01. The number of reads associated with this virus was significantly lower after bloom collapse, consistent with the drop in Microcystis-associated reads. Changes in reads associated with other viruses in our datasets were less pronounced. This suggests the Ma-LMM01-like virus is the best candidate for virus-mediated bloom collapse of those identified in this study.
Previous studies on Lake Mikata suggest that Ma-LMM01 infection is endemic within Microcystis blooms, while increased viral numbers correlate with non-microcystin producers outcompeting microcystin producers (Yoshida, et al. 2008; Yoshida, et al. 2010). However, instances of Ma-LMM01-mediated bloom collapse have not been identified. With the evidence presented here, we believe an Ma-L
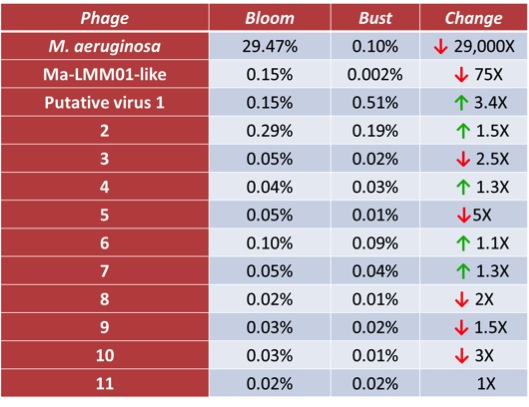
Table 1. Percent of total sequences from Bloom and Bust datasets aligned to assembled viral sequences, and their change in abundance from the Bloom to the Bust datasets.
MM01-like virus may be responsible for a Microcystis bloom collapse in the San Francisco Delta. We are investigating this further by using QPCR, which will quantify the ratio of virus to host (Microcystis cells) throughout this transect. This test will also allow us to track the presence and abundance of this virus in other freshwater systems.
I would like to thank WALPA and OLA for this poster award, and for hosting the Joint Lakes Conference this year. I also thank the Delta Stewardship Council for providing us the samples and environmental data we needed for this study.







